 |


|
Биология1966. Геодакян В.А., "Два пола. Зачем и почему? Эволюционная роль разделения на два пола с точки зрения кибернетики".Два пола. Зачем и почему?
Эволюционная роль разделения на два пола с точки зрения кибернетики. Геодакян В.А. Журнал «Наука и жизнь», 1966, № 3, стр. 99-105. А нужны ли самцы? В горах Армении, на берегах озера Севан, живут интересные ящерицы. Они замечательны тем, что среди них нет самцов – одни самки, которые откладывают неоплодотворенные, но способные к развитию яйца. Из них выводятся опять-таки исключительно самки. Такой метод размножения крайне прост и весьма рационален: потомство могут дать все особи, и отпадают трудности, связанные с поиском «супруга». Выходит в деле продолжение рода вполне можно обойтись без самцов. Другой интересный способ размножения встречается у обитателей наших озер – серебристых карасей. Они, так же как и ящерицы с берегов Севана, представлены одними самками. Но эти самки все-таки прибегают к услугам самцов... других видов рыб. Сперма «чужих» самцов активирует их икринки, побуждая их к развитию. Однако настоящего оплодотворения – слияния ядер мужской и женской половых клеток – здесь не происходит. Самцы в таком случае не принимают генетического участия в образовании новых существ и поэтому не могут претендовать на отцовство. Роль отца могут играть иголка или некоторые химические вещества. Например, икринки лягушки можно стимулировать к развитию уколом тонкой иглы, а яйцеклетки ряда морских животных побудить к делению, встряхивая их или добавляя к воде некоторые кислоты или соли. В лабораторных условиях даже столь высокоорганизованное животное, как кролик, иногда появляется на свет без участия самца. Яйцеклетку извлекают из организма самки, механическим или химическим путем побуждают к делению, а затем вновь помещают в чрево матери. Далее идет нормальное развитие и рождение нового существа. Такой «крольчонок-сирота» может развиться во вполне взрослое животное. У некоторых видов животных можно найти примеры весьма «несправедливого» отношения к самцам. Так самки пауков допускают к спариванию, но съедают его сразу после свадьбы. Чтобы избежать такой участи, самец преподносит своей кровожадной невесте лакомство. Также «безбожно» поступают и самки богомола, откусывающие во время спаривания голову самца. Свою миссию самцу-богомолу приходится выполнять, уже будучи без головы. К этим примерам можно добавить еще повадки пчел и ос, некоторые виды которых «допускают» появление самцов не в каждом поколении или практикуют их уничтожение сразу же после осеменения матки. Однако у подавляющего большинства животных самки «держат» своих самцов, терпят их и относятся к ним довольно сносно. Более того, виды, которые, «умеют» размножаться без самцов, например, некоторые ракообразные, обходятся без них летом – когда тепло и много корма. Но, как только наступает осень или засуха, они прибегают к услугам самцов. Это заставляет думать, что самцы для чего-то все-таки нужны. Попытаемся выяснить, для чего. Для чего нужно скрещивание? Существует два основных способа размножения: бесполое и половое. В бесполом размножении участвует только одна родительская особь, которая производит себе подобных. В половом размножении участвуют два родителя. Принципиальное значение имеет, однако, не количественная сторона: то, что в одном случае получается «из одного – два», а в другом – «из двух – три». Гораздо важнее качественная сторона дела, а именно то, что при бесполом варианте не возникает новых свойств, в то время как при половом каждый раз появляется иное качество, отличное от родительского. Это очень существенно. Бесполый способ встречается в основном у одноклеточных организмов, половым способом размножается большинство животных, в том числе млекопитающие. Видимо, с эволюционной точки зрения это наиболее прогрессивный способ. Преимущества полового размножения обычно связывают с возможностью перекомбинации наследственных признаков родителей, что приводит к появлению генетического разнообразия. В самом деле, легко родиться метисом, имея черного отца и белую мать. Но это невозможно без отца – только лишь от одной белой матери. Половое размножение обязательно предполагает скрещивание и, как правило, сопровождается разделением на два пола. Скрещивание – как раз тот процесс, благодаря которому возникают новые варианты. Оно необходимо для обеспечения генетического разнообразия. А что дает разделение на два пола? Нужны ли для скрещивания два пола? Существует способ размножения, при котором животные не разделены – не дифференцированы – на два пола, однако скрещивание имеет место. Таким способом размножаются дождевые черви. У них каждый червяк одновременно является и самкой и самцом. Или устрицы, у которых одна и та же особь выступает сначала в роли самца, а затем в роли самки. Казалось бы, такой способ дает больше преимуществ. Действительно, если, например, популяция животных состоит из 100 особей и все они без исключения могут скрещиваться между собой, то максимальное число возможных вариантов будет (100*99)/2 = 4950. Если та же популяция делится поровну на два пола, то число особей каждого пола равно 50 и максимальное количество возможных вариантов при этом равняется произведению 50*50 = 2500. То есть почти в два раза меньше. Как видим, разделение на два пола ухудшает дело. В чем же тогда секрет? Какую выгоду дает дифференциация полов? Ради чего популяция жертвует доброй половиной возможных вариантов, и что она получает взамен? Считается, что дифференциация на два пола обеспечивает производство двух сортов гамет – половых клеток: с одной стороны маленьких и подвижных сперматозоидов, задача которых состоит в том, чтобы встретиться с яйцеклеткой, и, с другой стороны, относительно больших, но неподвижных яйцеклеток, обеспечивающих будущий зародыш питательными веществами. Однако такая специализация имеет место и у гермафродитных животных (дождевых червей и устриц) без дифференциации на два пола и без уменьшения разнообразия вариантов. Следовательно, этим биологический смысл разделения на два пола объяснить нельзя. Попытаемся проанализировать роль полов в процессе воспроизводства, то есть выяснить их отношение к основным критериям производства – количеству, качеству и ассортименту продукции. Ибо всякое производство характеризуется прежде всего этими тремя параметрами. Количество, качество и ассортимент продукции. Предположим, что в заповедник выпускают всего 100 зубров. Какое выбрать соотношение полов, сколько выпустить коров и сколько быков? очевидно, что это соотношение зависит от того, какие цели преследуются. Если, например, задаются целью получить максимальное количество телят, то разумно взять 99 коров и 1 быка – при этом в каждом поколении могут родиться 99 телят. Однако все эти телята окажутся одного сорта по отцу и будут отличаться только по матери. Максимальное количество возможных сочетаний родителей в этом случае равно 99. Если желательно получить максимальное разнообразие (ассортимент), то необходимо иметь коров и быков поровну. Тогда количество возможных вариантов скрещиваний определяется произведением 50*50 = 2500! Однако уменьшится количество потомства, так как в одном поколении родиться лишь 50 телят. Среди них будут результаты как хороших сочетаний родителей, так и плохих. Если же желательно улучшить качество стада зубров, то необходимо создать условия для полового отбора, то есть сделать так, чтобы часть животных не участвовала в размножении. Для этого необходимо иметь избыток самцов. Тогда конкуренция в борьбе за самку приведет к тому, что часть самцов не будет представлена в потомстве. При этом, чем больше избыток самцов, те жестче будут условия отбора. Таким образом, существует определенная специализация – своего рода «разделение труда» - в роли полов в воспроизводстве. Оно выражается в их разном отношении к основным параметрам продукции – количеству и качеству потомства. Чем больше самок в популяции, тем больше количество потомства, чем больше самцов – тем лучше отбор, тем быстрее скорость изменения качества. Такая «асимметрия» возникает только на уровне популяции. (Речь идет об «идеальной», хаотически скрещивающейся популяции.) Каждому отпрыску отец и мать передают примерно одинаковое количество генетической информации. Если же рассматривать не семью, а популяцию в целом, появляются новые признаки, новые черты, свойственные данному, более высокому уровню организации. Сечение канала передачи генетической информации. Описанная специализация обусловлена тем, что потенциальные возможности самца в передаче генетической информации несравненно больше, чем возможности самки. Каждый самец в принципе может стать отцом всего потомства популяции, в то время как возможности самок в этом смысле ограничены. Говоря языком кибернетики, сечение канала связи самца с потомством значительно шире, чем сечение канала связи самки. Поэтому редкие варианты самцов в отличие от редких вариантов самок могут играть существенную роль в изменении среднего генотипа, то есть качества популяции. Разное сочетание каналов связи с потомством проявляется еще и в том, что каждый самец в отдельности «стремится» полнее использовать свои возможности и оставить максимальное количество потомства, влияя таким образом на качество популяции. В то же время каждая самка «стремится» к тому, чтобы то ограниченное количество потомства, которое она в состоянии оставить, было лучшего качества. Таким образом, если попытаться кратко, схематически сформулировать отношение полов к количеству и качеству потомства, можно сказать, что количество самок определяет количество потомства популяции, а каждая самка является «борцом» за качество потомства. Количество же самцов определяет качество популяции, а каждый самец является «борцом» за количество потомства. Эта упрощенная формула отражает также в какой-то степени тенденции различного поведения полов, их «психологию». Яркий пример такой разной «психологии» полов мы находим у Дарвина, который пишет, что «самцы оленьих гончих собак питают склонность к чужим самкам, тогда как самки предпочитают самцов, к обществу которых они привыкли». Это, конечно, не означает, что самки хорошие, а самцы плохие. Просто они разные, и это различие имеет биологическую основу. Для того, чтобы понять, какие выгоды дает виду специализация полов, необходимо рассмотреть взаимоотношения популяции со средой. Но прежде попытаемся выявить черты аналогичного строения различных управляемых систем с точки зрения кибернетики – науки об аналогиях. Что общего между ракетой, футбольной командой и популяцией животных? С точки зрения кибернетики это управляемые системы. Все они характеризуются движением к цели. Для ракеты этой целью может быть Луна, для футбольной команды – выигрыш, а для популяции животных – продолжение рода. Все три системы подвержены разным возмущающим помехам. Для ракеты это атмосфера или гравитационные поля, для футбольной команды – усилия противника, а для популяции животных – различные факторы среды: климат, пища, хищники, паразиты и прочие. Каждая система противопоставляет возмущающим помехам устойчивость своего движения. Каким образом достигается эта устойчивость? Обращает на себя внимание одна общая черта – разделение службы сохранения (консервативной службы), задача которой «сохранить все как есть», и службы изменения (оперативной службы) с задачей «ввести поправку». У ракеты службу сохранения осуществляют стабилизаторы, а службу изменения – рули. У футбольной команды это соответственно защита, которая стремится сохранить счет неизменным, и нападение, которое старается изменить его в пользу своей команды. Следовательно, один и тот же результат, устойчивость движения, в разных системах достигается аналогичным образом – разделением консервативной и оперативной служб. Такая дифференциация дает возможность системе добиться максимальной устойчивости движения. Как же обстоит дело в популяции животных? Не связана ли дифференциация на два пола с аналогичным выделением служб сохранения и изменения? Мы уже показали, что самки определяют количественную сторону размножения, а самцы – качественную. В биологических категориях это означает, что самки в популяции выражают в большей степени тенденцию наследственности, а самцы – изменчивости. И, наконец, применяя термины кибернетики, можно говорить, что самки олицетворяют «постоянную память» вида, в то время, как самцы олицетворяют «оперативную», «временную память» вида. Такая специализация – отделение оперативной памяти от постоянной – дает виду существенные преимущества. Чтобы убедиться в этом, перейдем к рассмотрению взаимоотношений популяции животных со средой. Фронт вредного фактора среды и кривая смертности. Понятие среды включает совокупность всех физических, химических и биотических факторов, с которыми организм сталкивается в течение своей жизни. Это прежде всего климатические факторы: холод и жара, повышенная влажность и засуха; это различные химические вещества в пище, в воде или в воздухе, и, наконец, это различные живые организмы того же или других видов, живущие в той же местности (хищники, паразиты, и прочие). Одно их характерных свойств живых систем – способность приспосабливаться к изменившимся условиям среды. Для этого система должна получать информацию от среды о происшедших изменениях. Все признаки организма прямо или косвенно связаны м соответствующими факторами среды: морозостойкость связана с низкими температурами, жаростойкость – с высокими, засухоустойчивость – с влажностью и т.д. Связь других признаков с условиями среды может быть менее очевидной, но не вызывает сомнений, что оптимальные, средние значения признаков в конечном счете определяются соответствующими факторами среды или их комплексом. Для простоты и наглядности возьмем одну пару фактор-признак: температуру и устойчивость к ней – и на этом примере рассмотрим связь популяции со средой. Если по оси абсцисс (Х) откладывать интенсивность вредного фактора среды – скажем, холода (или устойчивости к нему), а по оси ординат (Y) – количество особей, погибающих при определенных значениях его, то для популяции в целом получим какое-то распределение устойчивости, или «кривую смертности». Фронт вредного фактора среды – в нашем случае холода – можно представить в виде линии, которая отсекает самую чувствительную часть популяции (заштрихованная площадь на рис.1). Для того чтобы популяция «чувствовала» приближение фронта вредного фактора, необходимо, чтобы кривая смертности все время контактировала с этим фронтом, то есть за получаемую информацию, приводящую к изменению качества, популяция должна все время платить определенной жертвой количества.  Это означает, что даже в популяциях животных, обитающих в тропиках, обезьян, к примеру, какие-то экземпляры погибают от холода, в то время как в популяциях арктических или антарктических животных, например, пингвинов или белых медведей, какие-то особи погибают от жары. Такая плата за получаемую информацию является непременным условием связи со средой. Популяция, которая не платит регулярно этой дани, не получает информации от среды и лишена возможности приспосабливаться к ней. Резко изменившиеся условия среды могут застать такую популяцию врасплох и погубить ее. Естественно, популяции выгоднее свести к минимуму эту жертву количества за новое качество. Каким же образом достигается это? Прежде всего «асимметрия» отношения полов к количеству и качеству потомства приводит к тому, что воздействия среды на самок и самцов по-разному сказываются на характеристиках потомства. Потеря самок от неблагоприятных факторов среды сильно отражается на количестве потомства, не оказывая существенного влияния на его качество, тогда как потеря самцов в неблагоприятных условиях среды не отражается на количестве потомства и способствует изменению его качества в нужном направлении. Итак, можно сказать, что потеря самок от вредных условий среды приносит только вред, уменьшая численность популяции. Потеря же самцов приносит одновременно пользу, способствую эволюции вида. Является ли «прекрасный пол» слабым? Поэты и прозаики при определении женского пола, чаще всего пользуются эпитетами «прекрасный» или «слабый». Справедливость первого эпитета, кажется, не вызывает сомнений. Но соответствует ли истине второй эпитет? Если под силой понимать устойчивость, выносливость к различным неблагоприятным факторам, то сильным полом придется признать женский пол. Действительно, многочисленные эксперименты на растениях и животных и наблюдения на человеке показывают, что от всех губительных факторов среды: жары, холода, голода, различных ядов и болезней – прежде всего гибнут мужские организмы. При этом пониженной устойчивостью по сравнению с женским отличается не только мужской организм в целом, но также различные его органы, ткани и клетки. Чем же объясняется повышенная смертность мужского пола? Имеет ли это какой-либо биологический смысл? По этому вопросу существуют две теории. Согласно одной из них, смертность всегда выше у гетерогаметного пола вследствие наличия сцепленных с полом рецессивных летальных генов. (Напомним, что гетерогаметным называется пол с непарными половыми хромосомами – Х и Y, в противоположность гомогаметному полу, обладателю парных половых хромосом - Х и Х). Рецессивные летальные гены этих хромосом проявляются только у гетерогаметного пола. (По другой теории высокая смертность у самцов является результатом более интенсивного обмена веществ в их организме.) Против первой теории говорят результаты экспериментов по изучению смертности различных полов у птиц, бабочек и моли. У этих животных в отличие от подавляющего большинства остальных гетерогаметны самки, а гомогаметны самцы. В опытах было показано, что у многих бабочек, нескольких видов птиц и моли смертность самцов также почти всегда выше, чем смертность самок. Вторая теория фактически не объясняет ничего, заменяя непонятную «высокую смертность» самцов не менее непонятным «более интенсивным обменом веществ». Казалось бы, если существуют устойчивые формы самок, почему бы и самцам не обладать такой же устойчивостью? Повышенная смертность самцов... целесообразна. Итак, самцы являются биологически более слабым полом. Это означает следующее. Если мы нарисуем кривые смертности от данного вредного фактора отдельно для каждого пола, то с фронтом вредного фактора среды должна «контактировать» в основном кривая самцов. Следовательно, либо должен наблюдаться определенный сдвиг кривых смертности разных полов – таким образом, чтобы самцы оказались между фронтом вредного фактора среды и самками (рис.2), либо кривая смертности самцов должна обладать большей дисперсией (рис.3). 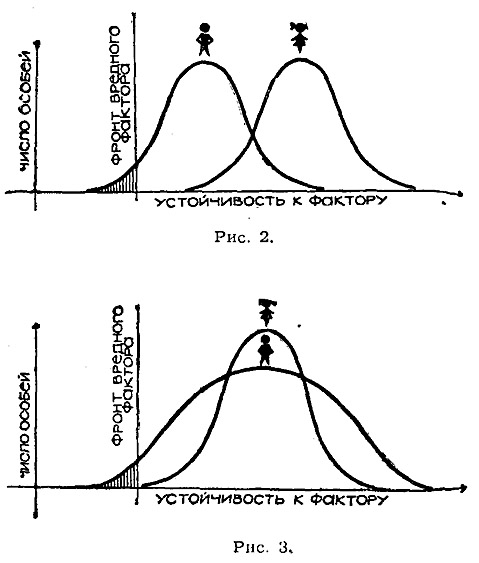 Сопоставляя эти соображения с выводами о том, что потеря самцов не влияет на количество потомства, но способствует изменению его качества в нужном направлении и что редкие варианты самцов имеют большее информационное значение, чем редкие варианты самок, то есть самцы являются основными переносчиками информации от среды к популяции, - приходим к заключению, что кривые распределения признаков для самцов должны отличаться от соответствующих кривых для самок большей дисперсией. Это значит, что самцы по всем признакам должны отличаться большим разнообразием, чем самки. Иначе говоря, если объединить всех самцов популяции в мужскую команду, а всех самок – в женскую и устроить личные и командные соревнования, то чемпионами в личном первенстве по всем видам программ будут самцы, а в командном зачете победят самки. Такие соотношения устойчивости дают возможность виду за новую информацию расплачиваться в основном самцами, потеря которых способствует сдвигу качества потомства, не отражаясь на его численности. Итак, более высокая смертность мужского пола целесообразна для сохранения вида. Связь соотношения полов при рождении с условиями среды. Из сказанного становится ясно, что соотношение полов является важным параметром популяции, тесно связанным с тенденциями наследственности и изменчивости в процессе воспроизводства. Следовательно, соотношение полов при рождении в разные периоды жизни популяции должно отражать эти тенденции в зависимости от условий среды. Повышенная гибель самцов в неблагоприятных условиях среды приводит в силу существования отрицательной обратной связи (о ней мы говорили в статье «Мальчик или девочка?». См. «Наука и жизнь» № 1, 1965 г.) к преимущественному рождению самцов. Увеличение рождаемости самцов может быть вызвано также непосредственно изменившимися условиями среды независимо от соотношения полов взрослых животных. Есть основания полагать, что у позвоночных такая регуляция осуществляется стероидными гормонами гипофиза, коры надпочечников и гонад. Следовательно, тяжелые условия среды должны повышать одновременно смертность и рождаемость самцов. Иными словами, можно сформулировать правило, согласно которому во всех неблагоприятных условиях среды должна увеличиваться «оборачиваемость» самцов, так как это способствует быстрейшему приспособлению вида. В биологической литературе можно найти массу фактов, подтверждающих эту закономерность При этом интересно, что виды, которые могут размножаться как бесполым способом, так и половым – бактерии, инфузории, некоторые ракообразные и другие, как правило, прибегают к половому размножению при наступлении неблагоприятных условий. Например, у многих водяных блох, а также тлей при благоприятных условиях – обычно летом – имеет место однополое размножение – так называемый партеногенез. Молодь (исключительно самки) вылупляется из «летних» яиц с мягкой оболочкой. При наступлении менее подходящих условий часть самок производит некоторое количество самцов, которые затем оплодотворяют самок. Оплодотворенные самки откладывают «зимние» яйца с твердой оболочкой, которые могут длительное время сохраняться в неблагоприятных условиях – на холоде, при большой жаре и пересыхании водоема (рис.4). 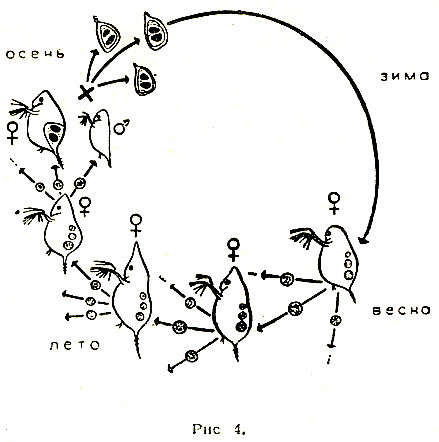 Перемещая коловраток из прудовой воды в речную или колодезную и обратно, ученые наблюдали на 3-й или 4-й день появление самцов, ранее отсутствовавших в популяции. При этом направление перемещения – происходило ли оно из прудовой воды в речную, или наоборот – оказывалось безразличным: любое изменение условий среды приводило к появлению самцов. Подвергая действию губительных факторов животных, размножающихся двуполым путем, например, плодовых мушек – дрозофил, ученые наблюдали одновременное повышение смертности и рождаемости самцов. Это казалось парадоксальным и необъяснимым. Действительно, почему одни и те же факторы – голод, холод, жара или яды – вызывают с одной стороны, гибель самцов, причем почти на всех стадиях развития, а с другой стороны – увеличение их числа в потомстве? Почему противоречивы порой теории, связанные с рождаемостью полов? Например, обратив внимание на то обстоятельство, что в холодный год рождается больше мальчиков, ученые делали вывод, что холод благоприятствует рождению мальчиков, а жара – девочек. Спустя некоторое время замечали, что сильная жара также сопровождается увеличением рождаемости мальчиков. Тут же возникала новая теория, согласно которой жара способствует появлению мальчиков, а холод – девочек. Между тем очевидно, что оба наблюдаемых явления легко объяснить с точки зрения правила увеличения «оборачиваемости» самцов при всех изменениях условий среды, требующих приспособления. Факты, подтверждающие это правило, можно найти и у млекопитающих, в том числе и у людей. Об этом говорят данные демографической и медицинской статистики, согласно которым во время больших климатических или социальных сдвигов (резкое изменение температуры, засуха, война, голод, переселения), то есть в периоды повышенной смертности, наблюдается тенденция роста процента мальчиков среди новорожденных. Ту же тенденцию отмечают и животноводы: чем лучше условия содержания сельскохозяйственных животных, тем больше самок в потомстве – даже при искусственном оплодотворении спермой одного и того же производителя. Разделение особей в популяции на два пола и «специализация» полов, при которой один из них «отвечает» за качество, а другой – за количество потомства, приводит к тому, что любой поток информации об изменении среды воспринимают сначала самцы, реагируют на него и преобразуют его. Иными словами, новая информация попадает сначала в «оперативную память» популяции, где подвергается проверке и отбору, и только после этого передается в «постоянную память» – самкам. Такое разделение на инерционное, стабильное ядро и более изменчивую, чувствительную оболочку, позволяет популяции отличить временные, короткодействующие и случайные изменения факторов среды, например, необычно холодную зиму, особенно жаркое лето, от систематических изменений в одном и том же направлении – скажем, наступления ледникового периода. Поэтому можно сказать, что поток информации из «оперативной памяти» в «постоянную» проходит через своего рода «частотный фильтр», который пропускает только малые частоты, задерживая большие. Инерционность «постоянной памяти» достигается именно этой задержкой новой информации в «оперативной памяти». Глина и мрамор. Прежде чем изваять статую из мрамора, хороший скульптор предварительно создает много вариантов из глины. Природа действует аналогичным образом, создавая большое разнообразие самцов, подвергая их проверке и отбирая необходимые варианты. Точно так же, как появление вариантов из глины всегда предшествует появлению совершенного варианта из мрамора, появление новых признаков у самцов должно предшествовать появлению тех же признаков у самок. Следовательно, мы можем рассматривать самцов как передовой отряд популяции, продвинутый далеко вперед, навстречу вредным факторам среды. Между этим авангардом и «ядром», «золотым фондом» популяции – самками – выдерживается определенная дистанция, необходимая для проверки и отбора. Эволюционная инерционность, отставание самок – это плата за их совершенство. И, наоборот, прогрессивность самцов – проявление их несовершенства. Эти представления можно сформулировать в виде гипотезы, согласно которой, все новые признаки в филогенезе должны закрепляться сначала у самцов, а потом передаваться самкам. Иными словами, можно сказать, что самцы являются «воротами» для изменчивости в наследственность. Следовательно, если самец и самка отличаются друг от друга по какому-то признаку (например, рост или цвет), то можно предсказать направление изменения этого признака. Оно будет совпадать с изменением от самки к самцу. Если самцы, например, крупнее самок, значит, существует тенденция укрупнения вида, если, наоборот, они мельче самок, - вид мельчает. Или, если самцы темнее самок, - окраска вида становится более темной и наоборот. У человека на данном этапе исторического развития можно предположить тенденцию к увеличению роста, поскольку средний рост мужчин выше, чем женщин (рис.5). У пауков, наоборот, наблюдается тенденция к уменьшению размеров, так как самцы у них мельче самок (рис.6). Антропологи и энтомологи считают, что это так и есть: человек растет, а паук уменьшается. 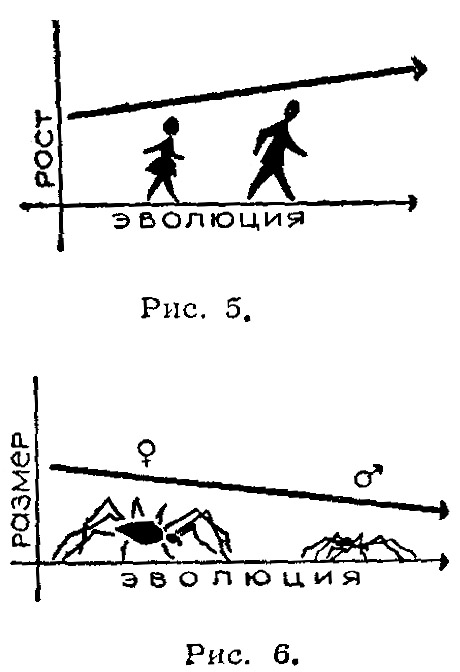 В качестве наглядного примера можно привести также известную картину онтогенетического и филогенетического появления рогов у самцов и самок оленей. Можно проследить четкую связь между степенью рогатости вида, с одной стороны, и возрастом появления рогов у самцов и самок – с другой. Чем сильнее выражена рогатость, тем в более раннем возрасте появляются рога сначала у самцов, а затем у самок. Очевидно, что предлагаемое правило можно применить как инструмент для исследования некоторых конкретных проблем эволюции. Не забывая, конечно, что эта общая тенденция может иногда перекрываться другими. Таким образом, применение общих идей и подходов кибернетики к постановке и решению проблем биологии позволяет понять некоторые непонятные до этого факты. Теперь мы знаем, что преимущества, даваемые однополым размножением, могут быть эффективны лишь на коротких, «спринтерских» дистанциях эволюции. «Стайерские» и «марафонские» дистанции можно пройти только раздельнополым способом. Ясно также, что дает скрещивание и что дает дифференциация, и то, что преимущества, обеспечиваемые ими, в полной мере проявляются только в «идеальной» популяции. В частности, с этим связан незначительный половой диморфизм моногамных видов и, наоборот, четко выраженный – полигамных. В заключение, возвращаясь к вопросу, нужны ли вообще самцы, можно ответить: да, нужны! Они нужны в основном для приспособления к изменившимся условиям среды. Это справедливо для животных. А как же для человека? Ведь известно, что ускоренные темпы социальной и технической эволюции неуклонно уменьшают роль биологической эволюции. Научившись изменять условия среды, человек тем самым освобождается от необходимости изменяться самому. В самом деле, если вновь наступит холодный, ледниковый период, животные отрастят густую шерсть, а человек наденет синтетическую шубу. Таким образом, можно предположить, что социальный и технический прогресс будет сопровождаться неуклонным возрастанием роли и доли женщин в обществе. Справка: Геодакян Виген Артаваздович, кандидат технических наук (физико-химик) и доктор биологических наук (биолог теоретик), работал в ФИАН, институтах Молекулярной биологии, Биофизики, Общей генетики, Биологии развития, Институте человека. С 1990 г. ведущий научный сотрудник Лаборатории биоакустики Института эволюционной морфологии и экологии животных РАН им. А.Н. Северцова (Москва). Автор эволюционных теорий пола, полового диморфизма, гоносом и ”номадических” генов, асимметрии организмов, функциональной асимметрии мозга, парных органов и др. Научные интересы - связанные с полом проблемы эволюции, генетики, экологии, асимметрии мозга и психологии, а также вопросы информации и организации систем. E-mail: geod@newtech.ru Геодакян В.А. Роль полов в передаче и преобразовании генетической информации // Проблемы передачи информации. 1965. Т. 1. № 1. С. 105-112. Геодакян В.А. Дифференциальная смертность и норма реакции мужского и женского пола // Журн. общ. биол. 1974. Т. 35. № 3. Геодакян В.А. Эволюционная логика дифференциации полов // Природа. 1983. № 1. С. 70-80. Геодакян В.А. Онтогенетическое правило полового диморфизма // Докл. АН СССР. 1983. Т. 269. № 12. С. 477-482. Геодакян В.А. О теоретической биологии / Методологические аспекты эволюционного учения. Киев, 1986. Геодакян В.А. Теория дифференциации полов в проблемах человека // Человек в системе наук. М., 1989. С. 171-189. Геодакян В.А. Эволюционная теория пола // Природа. 1991. № 8. http://vivovoco.rsl.ru/VV/PAPERS/NATURE/VV_SC30W.HTM Геодакян В.А. Два пола: зачем и почему? СПб., 1992. |
| © 2005 - 2009 Е.В. Вейник работает на Sitefactor.CMS |
|